
懷孕需要生理性暴露於外來胎兒同種異體抗原,而且經常會再次暴露於異源胎兒同種抗原。懷孕後的後果差異很大,有同種免疫和耐受性表型增加的證據。我們表明,懷孕引發了胎兒特異性母體CD8 + T細胞的積累及其在分娩後作為活化記憶池的持久性。在非生殖環境中,抗原再相遇會發生細胞溶解和強大的二次擴增潛能。相比之下,與PD-1和LAG-3表達增加相關的CD8 + T細胞功能衰竭與隨後懷孕期間胎兒抗原的重新刺激有關。PD-L1 / LAG-3中和釋放了胎兒特異性CD8 +的活化T細胞,在繼發而非原發妊娠期間選擇性地導致胎兒浪費。
為了更全面地研究母親在懷孕期間對胎兒刺激的免疫學反應,將主要的組織相容性複合體(MHC)I類四聚體染色與四聚體陽性細胞富集相結合,以鑑定具有確定胎兒特異性的罕見內源性母體CD8 + T細胞。該方法利用了跟踪內源性母體T細胞的優勢,但同時繞過了僅對一部分外周T細胞進行採樣所帶來的限制。通過重新定位這些工具,可以識別出分娩後仍然存在的由妊娠引發的胎兒特異性CD8 + T細胞的獨特庫,並證明了其上下文特異性同種免疫與耐受性表型相關的分子基礎。
妊娠後具有胎兒特異性的母體CD8 + T細胞持續存在
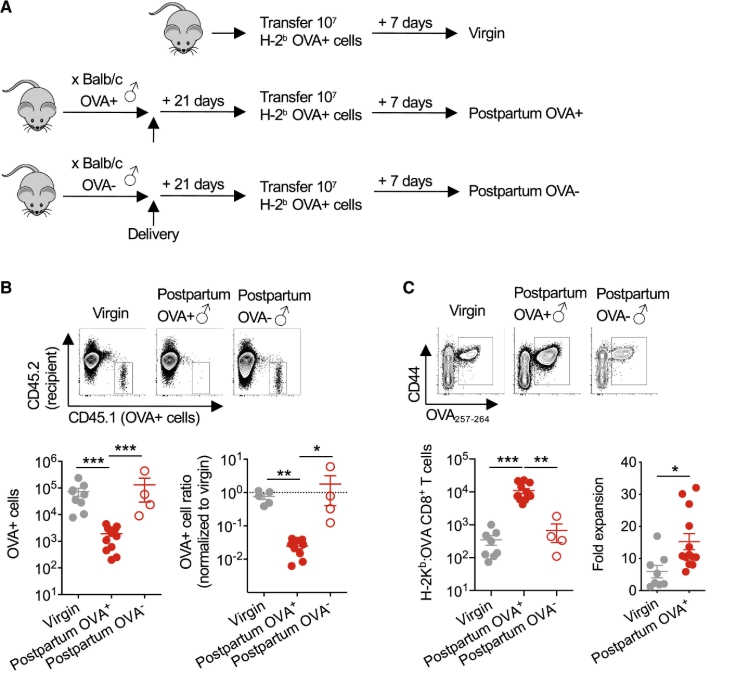
鑑於MHC單倍型等位基因中定義抗原的可用性有限,具有組成性細胞表面OVA表達的BALB / c(H-2 d)雄性小鼠被用於在非轉基因C57BL / 6(H-2 b)雌性中進行同種異體妊娠(圖1 A)。這種交配方案將OVA轉化為替代胎兒抗原,並允許使用MHC I類四聚體染色和富集來鑑定內源性,多克隆胎兒特異性(H-2K b:OVA 257-264)CD8 + T細胞。使用這種方法,我們顯示懷孕在脾臟和周圍淋巴結的妊娠中期(胚胎日[E] 10.5)啟動了具有胎兒OVA特異性的母體CD8 + T細胞的約50倍擴增(圖1 B) 。與原始對照小鼠中具有相同特異性的細胞相比,這些細胞的數目在分娩後收縮,直到分娩後8週仍保持> 10倍的增加水平(圖1 B)。
還觀察到由胎兒抗原刺激引發的母體CD8 + T細胞的表型轉移。在妊娠和分娩後,具有胎兒-OVA特異性的母體CD8 + T細胞顯示出更高的親和力,基於H-2K b:OVA四聚體染色強度和歸一化至TCR表達水平後的親和力,與原始對照中具有相同特異性的細胞相比小鼠(圖 1C)。懷孕期間和分娩後第21天,大多數胎兒OVA特異性CD8 + T細胞均為CD44 hi,表明有抗原經歷(圖1D)。然而,CD62L的持續下調以及CCR7表達的增加模糊了進一步分類為中央或效應記憶細胞的作用(圖1 D)。孕育的孕產婦CD8 + T細胞也顯示出明確的陰性共刺激分子(包括PD-1和LAG-3)表達增加,而在分娩後表達仍保持升高,而其他表達,包括CTLA-4和TIM-3,懷孕期間沒有明顯變化,或者分娩後沒有持續變化(圖1 D)。因此,原發妊娠會引發胎兒特異性CD8 +的擴增 T細胞作為獨特的記憶庫而持久,並具有與活化和功能衰竭相關的分子的豐富表達。
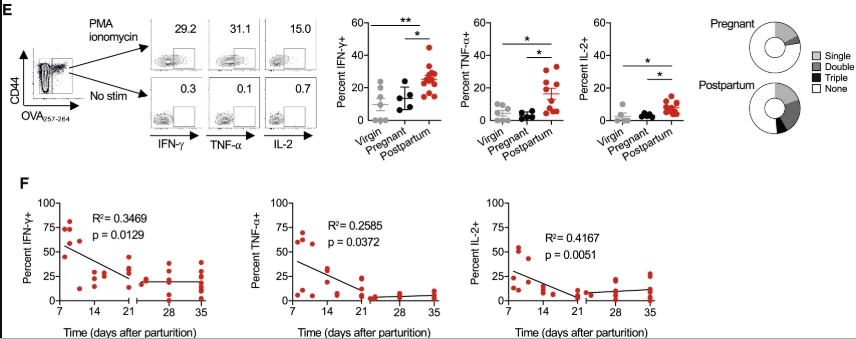
具有胎兒特異性的母體CD8 + T細胞在妊娠期間擴增並在分娩後持續存在。(圖源:Sciencedirect)
母體記憶CD8 + T細胞具有細胞溶解能力和效應細胞因子的產生
為了進一步研究胎兒OVA特異性CD8 + T細胞的功能特性,評估了它們產生細胞因子的潛力。生產規範效應細胞因子干擾素γ(IFN-γ),腫瘤壞死因子α(TNF-α)和白細胞介素-2(IL-2)H-2K b:OVA CD8 +以下佛波醇12-荳蔻酸酯13- T細胞與原始或妊娠中期妊娠小鼠中具有相同特異性的細胞相比,分娩後第21天時乙酸-乙酸(PMA)-離子黴素的刺激顯著增加(圖1 E)。孕婦CD8 +與胎儿期間具有相同特異性的細胞相比,具有胎兒OVA特異性的T細胞在分娩後同時產生多種效應細胞因子方面也表現出更高的多功能性(圖1 E)。額外的時程分析顯示,分娩後早期細胞因子的產生甚至更高,在分娩後第21天穩定下來(圖1 F)。因此,鑑於在這個時間點記憶細胞的功能穩定,後續實驗著重於產後第21天。
具有胎兒特異性的母體CD8 + T細胞在妊娠期間擴增並在分娩後持續存在。(圖源:Sciencedirect)
為了更準確地說明母親如何對完整的胎兒細胞進行免疫反應,評估了對來自Act-OVA轉基因小鼠的過繼轉移脾細胞的持久性和CD8 + T細胞反應(圖2 A)。我們發現,表達OVA的雄性與先前懷孕有關的雌性中,OVA +細胞的清除速度明顯加快(圖2 B)。轉移後第7天,與初次控制的小鼠相比,在先前懷孕的母親中,表達OVA的雄性導致OVA +(CD45.1 +)細胞減少了約100倍(圖2 B)。加速拒絕OVA +這些細胞與表達OVA的雄性有先孕的小鼠中胎兒OVA特異性CD8 + T細胞積累的增加平行(圖2 C)。這些差異反映了加速的擴增,並且不能解釋為與OVA +細胞轉移之前的原始小鼠相比,產後小鼠的OVA特異性CD8 + T細胞數量增加,因為與原始小鼠相比,產後的倍數擴展明顯增加(圖2 C)。
妊娠為首的CD8 + T細胞具有細胞溶解作用,並具有強大的繼發性再增殖能力(圖源:Sciencedirect)
繼發妊娠期間的母體記憶CD8 + T細胞低反應性
先前懷孕的這些保護性益處以伴侶特異性的方式發生,這表明,儘管懷孕引發了對胎兒抗原的同種免疫,但在隨後的懷孕期間,致耐受性反應得以恢復甚至增強。在非生殖環境中評估胎兒alloantigen反應時,為什麼致敏性占主導地位的一種解釋是,需要特定於妊娠的生理變化才能刺激孕前孕育的孕產婦免疫成分的低反應性表型。
通過比較記憶CD8 + T細胞中這些功能的分子基礎,通過比較其細胞內力消耗分子的表達與繼發妊娠期間胎兒抗原的再刺激來進行比較。與繼發妊娠期間胎兒OVA重新刺激導致CD8 + T細胞的鈍性擴張相一致,這些細胞顯示PD-1,LAG-3和CTLA-4共抑制分子的表達急劇增加,而TIM-3保持不變。
PD-1和LAG-3 在繼發妊娠期間選擇性損害CD8 + T細胞活化
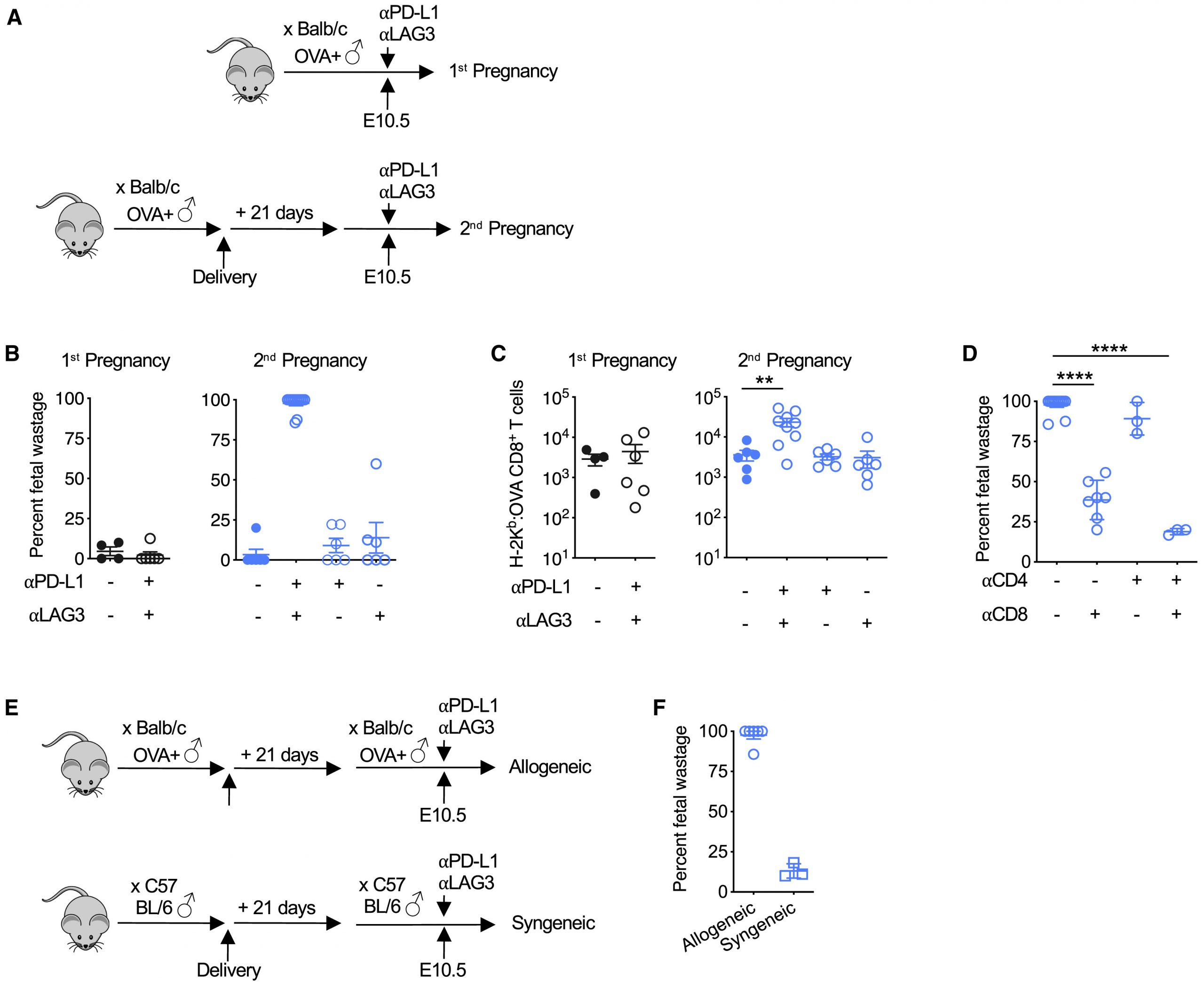
為了研究繼發妊娠期間母體T細胞選擇性擴增PD-1和LAG-3表達的功能意義,評估了中和這些分子對妊娠結局和胎兒特異性CD8 + T細胞積累的影響(圖5 A )。在繼發而非初次妊娠期間的妊娠中期給予PD-L1和LAG-3中和抗體會在72小時內引發幾乎完全的胎兒流失(圖5 B)。
PD-L1 / LAG-3功能性封鎖引起的胎兒浪費與繼發妊娠期間胎兒OVA特異性CD8 + T細胞的釋放重新釋放有關,但對初次妊娠期間這些細胞的積累沒有顯著影響(圖 5C)。同時中和PD-L1和LAG-3是必需的,因為在二次妊娠期間,僅用抗PD-L1或LAG的中和抗體治療小鼠時,既不會浪費胎兒,也不會發生具有胎兒OVA特異性的CD8 + T細胞的再擴增。-3(圖5 B和5C)。
PD-1和LAG-3 在繼發妊娠期間選擇性限制胎兒特異性CD8 + T細胞的再生。(圖源:Sciencedirect)
繼發妊娠期間由PD-L1 / LAG-3阻滯觸發的胎兒浪費直接反映了胎兒特異性CD8 + T細胞的釋放活化,因為隨著母體CD8 + T細胞與PD-L1 的共同消耗,胎兒浪費顯著減少/ LAG-3功能封鎖(圖5 D)。
這些妊娠結局表型也無法通過潛在的PD-L1 / PD-1或LAG-3直接刺激CD4 + T細胞來解釋,因為這些細胞的耗竭都不會推翻CD8中PD-L1 / LAG-3阻滯誘導的胎兒浪費+ T細胞充足的小鼠,或CD8 + T細胞枯竭的小鼠中對胎兒浪費的保護(圖5)D)。
然而,由PD-L1 / LAG-3阻斷觸發的胎兒浪費在MHC相同的雄性和雌性小鼠繼發的同基因妊娠中被推翻,這表明母體對遺傳上異源的胎兒同種異體抗原的識別驅動了CD8 + T細胞功能衰竭的必要性防止胎兒流失(圖5 E和5F)。
因此,與原代同種異體妊娠相比,繼發性胎兒刺激下的母體CD8 + T細胞表達的PD-1和LAG-3水平逐漸升高,並且這些共抑制分子通過抑制母體的記憶活化來防止胎兒浪費CD8 +T細胞是由先前的懷孕引發的。這些結果突出了與初次妊娠相比,初次妊娠和胎兒T細胞對胎兒抗原刺激反應如何達到耐受性的獨特策略。
結果
懷孕仍然是免疫學上的奇蹟和悖論。儘管有許多機制可以保護同種異體胎兒組織免於過早排斥,但是我們關於妊娠如何在免疫學上起作用的核心知識仍然是基本知識。在這方面,可以通過研究均等來發現重要的線索,以及與活化的免疫細胞相比,幼稚與胎兒抗原刺激反應的潛在差異。
與對外源性微生物入侵者俱有特異性的保護性記憶免疫細胞的加速活化和擴增相一致,胎盤哺乳動物也可能發展了目的論上保守的途徑,以記住有益的胎兒同種異體抗原並在隨後的暴露過程中增強胎兒的耐受性。
參考資料:
CD8+ T Cell Functional Exhaustion Overrides Pregnancy-Induced Fetal Antigen Alloimmunization